What Are Organic Molecules?
Organic molecules contain carbon-hydrogen bonds, are found in living things and can be very large molecules. The major classes of organic macromolecules are carbohydrates, proteins, lipids and nucleic acids.
What Are Nucleotides really?
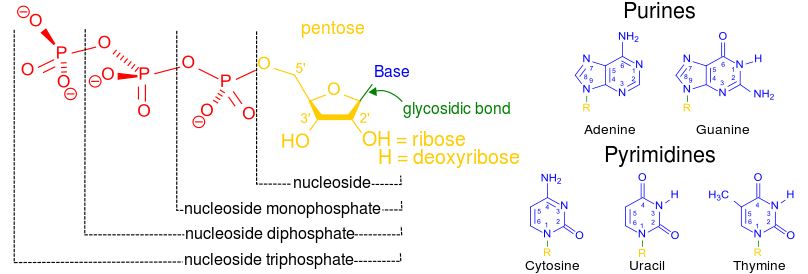
Nucleotides are monomers (small molecules) that are the building blocks of nucleic acids. Each nucleotide, and consists of 3 portions:
-a pentose sugar called ribose
-one or more phosphate groups
-one of five cyclic nitrogenous bases
Some nucleotides are put together to form nucleic acid (DNA & RNA) macromolecules, whereas others function on their own.
Nucleic Acid Structure
Nucleotides can be linked together by covalent bonds between the phosphate of one nucleotide and the sugar of next. These linked monomers become the phosphate-sugar backbone of nucleic acids. The nitrogenous bases extend out from this phosphate-sugar backbone like teeth of a comb.
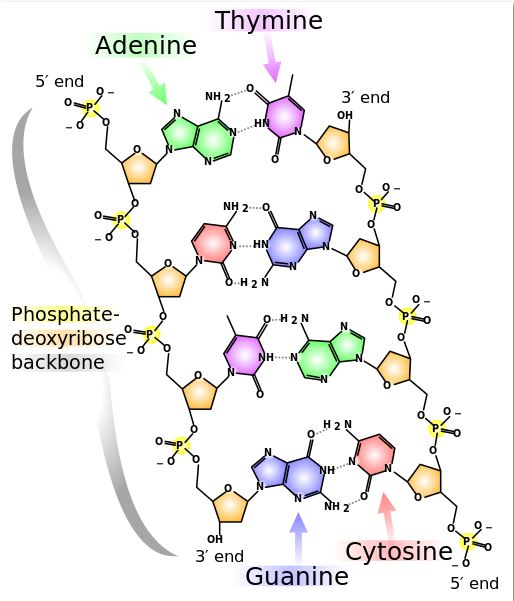
Deoxyribonucleic Acid (DNA)
DNA (deoxyribonucleic acid) is the genetic material, the original blueprint, inside each biological cell. The molecule is double-stranded and twisted, like a spiral staircase, with the two sugar-phosphate chains as the side rails, and the nitrogenous base pairs, linked by hydrogen bonds, forming the rungs. In addition to linking the bases together, hydrogen bonding twists the phosphate-sugar backbones into a helix, thus DNA is a double helix.There are four different types of nitrogenous bases that can be found in a DNA molecule: adenine (A), guanine (G), cytosine (C) and thymine (T). Adenine and guanine are larger, double ring nitrogenous bases called purines. Cytosine and thymine are smaller, single ring nitrogenous bases called pyrimidines. When bases pair up between the two DNA strands, a purine always pairs with a pyrimidine. Specifically adenine (A) and thymine (T) pair up, and cytosine (C) and guanine (G) pair up. These bases are attracted to each other through hydrogen bonding.When the DNA molecule is inactive, the bases are linked by these hydrogen bonds and the molecule is in its spiral-shaped state. When DNA is being used—either being copied (a process called replication) or being employed to build proteins (involving the processes of transcription and translation)—the DNA molecule must be opened up, essentially “unzipped” between the bases.
Ribonucleic Acids (RNA)
In living organisms, RNA is a single stranded nucleic acid molecule. In viruses, non-living infectious particles, RNA can be single or double stranded.There are four different types of nitrogenous bases found in an RNA molecule: adenine (A), guanine (G), cytosine (C) and uracil (U). In RNA, uracil takes the place of the thymine found in DNA.
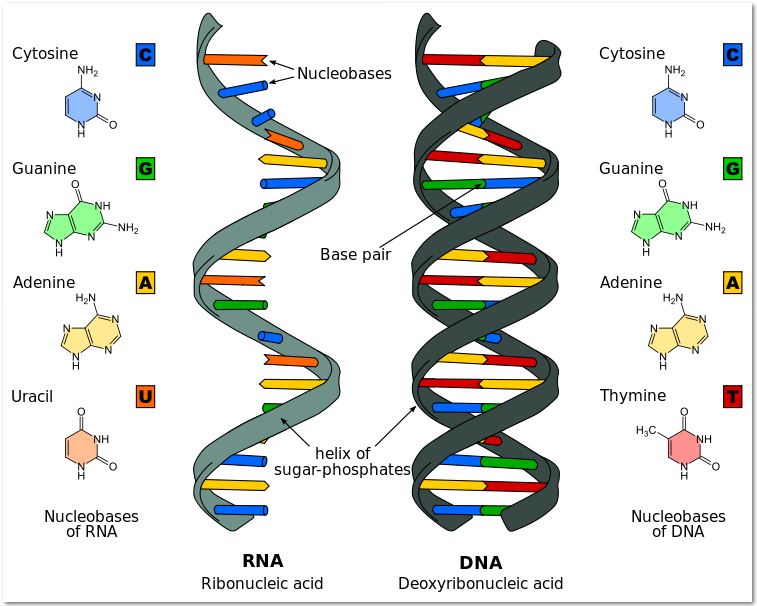
When RNA bases are laid down to build an RNA molecule, DNA is unzipped, and the new RNA molecule made is compliment of the DNA template. For example, if the DNA strand has the following bases, in this order, ATTGCACT, the new RNA molecule being made will have the base sequence UAACGTGA. After the RNA segment is made, the DNA zips back up and the RNA floats off to carry out its function in the cell.Genetic information copied from DNA is used to build three types of RNA:
1) Ribosomal RNA – The Protein Factories: Most of the RNA in cells is part of the structure of small cellular organelles known as ribosomes, the protein factories of the cells.
2) Messenger RNA – The Genetic Blueprint: Messenger RNA is a copy of the genetic information that was transcribed from the cell’s original blueprint, DNA. This copy of the genetic information is brought to the ribosome and used as instructions for building proteins.
3) Transfer RNA – The Amino Acid Suppliers: Transfer RNA is also part of the process of building proteins. Like a little truck, tRNA brings the amino acid to the ribosome. Which amino acid it brings depends on which was coded for in the mRNA instructions. At the ribosome, these amino acids are joined together to form proteins.
ATP: The Energy Transfer Molecule
Adenosine 5′-triphosphate (ATP) is a multifunctional nucleotide, most important as the “molecular currency” of intracellular energy transfer. Like tiny rechargeable batteries, ATP molecules transport chemical energy within a biological cell. These molecules can move energy around because the phosphate bonds contain a lot of potential energy, which is released when they are broken.During photosynthesis and cellular respiration, ATP is produced from ADP (adenosine diphosphate), an inorganic phosphate and added energy. ATP energy is consumed by a multitude of cellular processes.

So how was my presentation:

reference:
“What are Nucleotides and Nucleic Acids,” Science Prof Online; accessed on April 13, 2013.http://www.scienceprofonline.org/chemistry/nucleotides-nucleic-acids-atp-rna-dna.html








{kind=link}